摘要:
本研究分别进行了消化率试验和生长试验。试验1测定肉骨粉、猪肉粉、水解羽毛粉、鸡肉粉、鱼粉、豆粕和喷雾干燥血粉等7种原料的营养素和能量表观消化率。试验2为8周的生长试验,在可消化理想蛋白条件下设定两个蛋白水平,评估猪肉粉替代鱼粉的可行性。鱼粉组(FM组)为对照组(鱼粉:320g/kg,粗蛋白:434.9g/kg,粗脂肪:124.6g/kg),其它三个试验组用115g/kg猪肉粉替代160g/kg鱼粉,其中在与对照组同一粗蛋白水平中添加必需氨基酸组为PMA组,不添加组为PM组;LPMA组蛋白水平降低(粗蛋白:400.9g/kg,粗脂肪:96.3g/kg),但可消化蛋白能比和必需氨基酸模式与FM组相同。试验结果表明:各原料的蛋白消化率为85.6%~95.5%,氨基酸消化率为87.6%~95.9%。鱼粉和猪肉粉的蛋白表观消化率分别为91.2%和95.9%。在生长试验中,PMA组鱼的增重率和饲料系数与对照组无显著差异,但显著高于PM和LPMA组(P<0.05)。生长与赖氨酸和蛋氨酸摄入量呈线性关系。本试验结果表明,在理想蛋白条件下,可以用115g/kg的猪肉粉替代饲料中160g/kg的鱼粉。必需氨基酸缺乏(PM组)或在理想氨基酸模型条件下降低饲料蛋白水平(LPMA组),都不能满足日本海鲈达到最适生长的需求。
关键词:日本海鲈、消化率、理想蛋白、蛋白水平、鱼粉、猪肉粉
1.前言
日本海鲈是一种肉食性鱼类,在中国沿海以及东南亚各国广泛养殖(Xue等,2006)。肉食性海水鱼类饲料中鱼粉含量较高,通常在300~600g/kg之间,因此肉食性海水鱼饲料价格都较昂贵(Wang等,2006)。要提高海水鱼料的经济效益,不仅需要我们准确了解商业蛋白源的营养素消化率,饲料配方也需要进一步研究优化。目前为止已经有大量的研究关注于降低饲料中鱼粉的比例,有些研究报道显示不添加鱼粉或添加极少量鱼粉的饲料能够被肉食性鱼类很好的利用(Kaushik等,2004;Coves等,2005)。但是肉食性鱼类对低效价的替代蛋白源利用能力差异很大(Bureau等,2000;Milliamena,2002;Gómez-Requeni等,2004;Webster等,2004;Gaylord等,2005;Wang等,2006;Wang等,2008;Figueiredo-silva等,2010)。
在理想氨基酸模式下,饲料中最适的蛋白水平是鱼类维持正常生长和氮保留的先决条件(Luo等,2009)。测定营养素的消化率是评估水产饲料原料利用潜能的首要步骤(Allan等,2000)。与饲料中蛋白源鱼粉相比,限制其它饲料替代蛋白源利用的最重要因素是必需氨基酸的缺乏(Wilson,2002)。为了更加精确地制作饲料配方且预测鱼体的生长状况,同时参考氨基酸的最佳需要量和营养物质的消化率要比只关注饲料的总的氨基酸含量更加重要,这一理念已经在畜禽动物中成功应用(Dari等,2005;Libao-Mercado等,2006)。本试验的研究目的是测定几种饲料原料的表观消化率以及在日本海鲈两种蛋白质水平下肉骨粉部分替代鱼粉的研究。
2.材料与方法
2.1饲料制作
试验1,测定日本海鲈对鱼粉、牛肉骨粉、猪肉粉、水解羽毛粉、鸡肉粉、豆粕和喷雾干燥血粉中干物质、氮、总能、总磷和氨基酸的表观消化率。
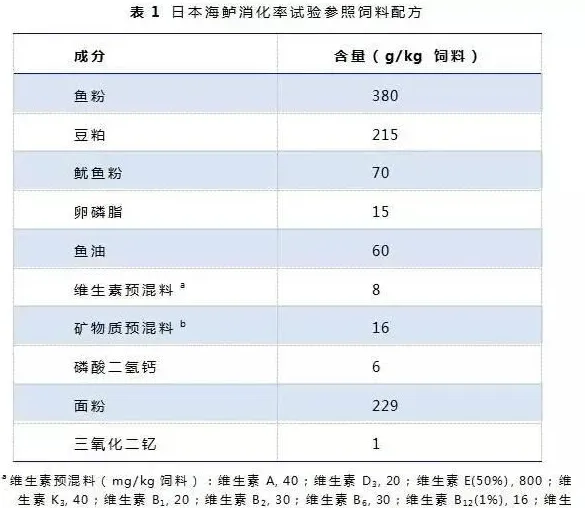
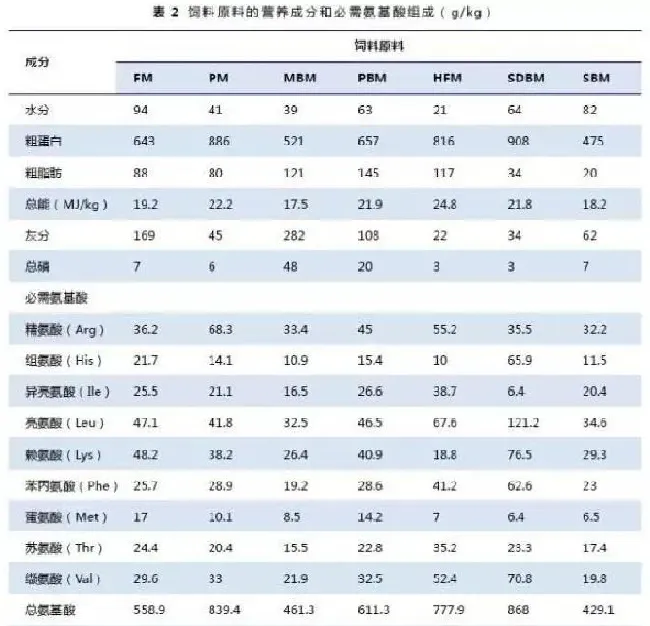
设置一组参照饲料,参照饲料与待测原料按70:30的比例混合,配制成另外7种试验饲料(表1)。三氧化二钇(Y2O3,1g/kg)作为内标物添加到参照饲料和试验饲料中。饲料原料营养成分和氨基酸组成见表2。
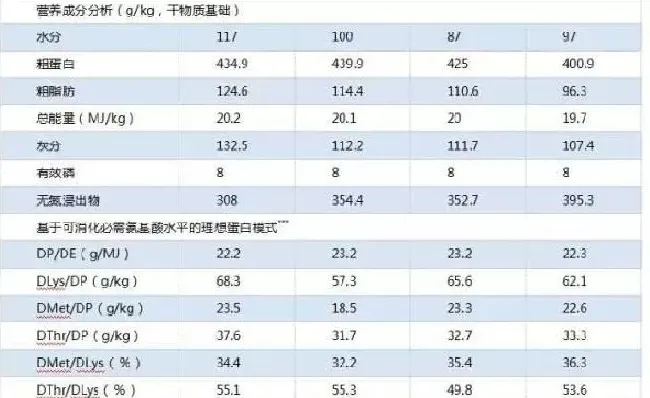
试验2,配制4种试验饲料,进行为期8周的养殖试验(表3)。鱼粉组为对照组(FM组,鱼粉:320g/kg,粗蛋白:434.9g/kg,粗脂肪:124.6g/kg,Ai等,2004),能满足日本海鲈对蛋白质和必需氨基酸的需要。其它三组均添加115g/kg的猪肉粉和160g/kg的鱼粉的饲料。PMA组和PM组与对照组粗蛋白水平相同,其中PMA组添加必需氨基酸,PM组不添加。LPMA组的蛋白水平降低(粗蛋白:400.9g/kg,粗脂肪:96.3g/kg)可消化蛋能比和必需氨基酸模式与FM组相同。实验2的赖氨酸、蛋氨酸和苏氨酸都达到需要量水平。日本海鲈的赖氨酸需要量为60.7±3.0g/kg饲料蛋白(Mai等,2006)。蛋氨酸和苏氨酸的需要量根据日本海鲈的肌肉氨基酸模型调整为21.8和30.4g/kg饲料蛋白(Mai等,2006)。饲料中有效磷水平用磷酸二氢钙调平衡(Zhang等,2006)。
饲料原料过60目筛,充分混匀后采用双螺杆挤压机(TSE65;北京洋工机械,中国)制成2mm直径的膨化料,经风干后4℃贮藏。挤压机筒温度(进料段—加压段—出料段):90℃5秒—130℃3秒—60℃4秒。
2.2饲养管理
日本海鲈来自山东潍坊渔场,用对照组饲料在循环系统中驯养2周。试验1开始前停喂一天,每个圆锥缸(直径80cm,体积0.25m3)中随机放入35尾初始重(26.0±0.3g)一致的鱼,每组饲料随机分配3个缸,共24个缸,养殖6周。根据试验1所得消化率数据,设计生长试验(试验2),使用同一批次鱼苗。将初始重60.0±0.1g的日本海鲈随机分配到16个圆锥缸中(直径80cm,体积0.25m3),每组饲料4个重复,每个重复25尾鱼。试验2结束后,每缸取2尾鱼分析全鱼组分,5尾鱼做形态学指标和血液学指标的测定。从鱼尾静脉采血后,血液3×105g、4℃离心10min,取上清-80℃保存。
饲养期间,每天投喂2次(上午9:00,下午16:00),温度维持在25~28℃,pH8.2~8.6,氨氮<0.4mg/L,亚硝酸盐<0.1mg/L,溶氧>6.0mg/L。各缸每天24h曝气,光周期为12h暗:12h亮,光照强度为7光照度(lx)。
2.3粪便收集
试验正式开始一周后开始收集粪便,以保证有足够的时间清除之前食物留下的肠内容物。使用能够将粪便和流水分开的沉降柱,为了使得粪便中的营养物质流失最小,只收集鱼体摄食1h后完整的粪便。每天将同一组的粪便样品合并,直到收集到足够的样品用于化学成分分析。粪便保存在-20℃。
2.4血液学指标和化学成分分析
测定所有原料、饲料、粪便和全鱼样品的干物质、粗蛋白、脂肪、灰分和能量,以及所有原料、饲料和粪便中的氨基酸含量。用自动分析仪和试剂盒测定血清中谷丙转氨酶、谷草转氨酶、甘油三酯、总胆固醇、葡萄糖和尿素氮。
干物质测定采用105℃干燥恒重法;粗蛋白测定采用凯氏定氮法(Thiex等,2002);利用酸水解索氏抽提法测定样品的脂肪含量(Foss,HillerØd,Denmark);灰分测定采用550℃灼烧法;总能测定采用PARR1281自动氧弹式量热计(Parr,Moline,IL,USA);利用全自动氨基酸分析仪(Hitachi8800,Tokyo,Japan)测定氨基酸含量;利用电感耦合等离子体原子发射光谱法(ICP-OES,JY38S,JobinYvon,ChillyMazarin,France)测定饲料、粪便中内标物三氧化二钇(Y2O3)和总磷的含量。
2.5消化率的计算及统计分析
参照饲料和试验饲料的干物质、粗蛋白、总能、总磷和氨基酸的表观消化率(ADC)的计算参照以下公式:
饲料营养素和总能ADC(%)=100×[1-(粪便营养素或总能含量/饲料中营养素或总能含量)×(饲料中Y2O3含量/粪便中Y2O3含量)]
原料ADC参照Bureau,Harris&Cho(1999)的计算方法:
原料ADC(%)=ADCTD+[(ADCTD-ADCRD)×(0.7×NutrRD/0.3×NutrING)]
ADCTD:试验饲料中营养素或能量的表观消化率
ADCRD:参照饲料中营养素或能量的表观消化率
NutrRD:参照饲料中营养素或能量的含量
NutrING:试验原料中营养素或能量的含量
应用STATISTICA6.0软件对数据进行单因素方差分析,差异显著者进行Duncan氏法多重比较,显著水平为P<0.05。
3结果
3.1原料表观消化率
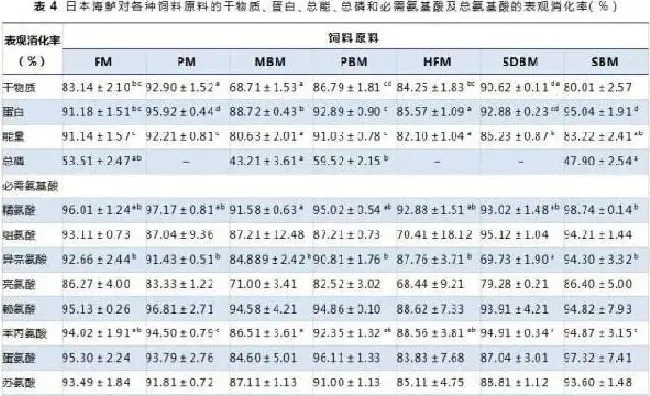
试验1和试验2各组的鱼存活率均很高(>98%)。日本海鲈对各种饲料原料的干物质、蛋白、总能和总磷表观消化率见表4。干物质表观消化率从牛肉骨粉的68.7%到猪肉粉的92.9%。营养素的表观消化率在各原料中显著不同。蛋白表观消化率在猪肉粉和豆粕中最高(P<0.05),其它原料的蛋白表观消化率也高于85.6%。猪肉粉的能量表观消化率显著高于牛肉骨粉、水解羽毛粉、喷雾干燥血粉和豆粕(P<0.05)。鱼粉、牛肉骨粉、鸡肉粉和豆粕的总磷表观消化率为43.2%~59.5%,其中牛肉骨粉的最低。另外,由于猪肉粉、水解羽毛粉和喷雾干燥血粉中的磷含量非常低,导致这几种原料的磷表观消化率计算值不可靠,因而被剔除掉。各原料中的氨基酸表观消化率不同,猪肉粉和鱼粉的氨基酸表观消化率相似,因而猪肉粉可作为鱼粉替代的蛋白源。
3.2试验2的生长指标
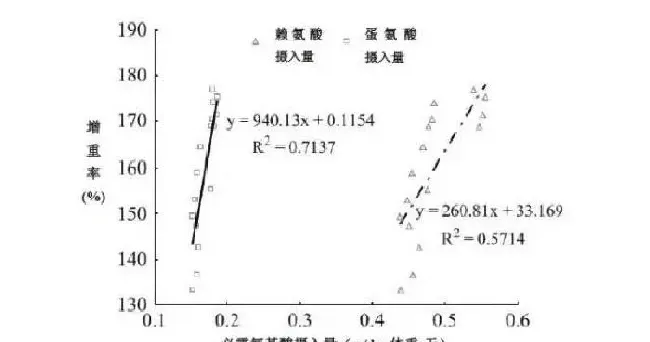
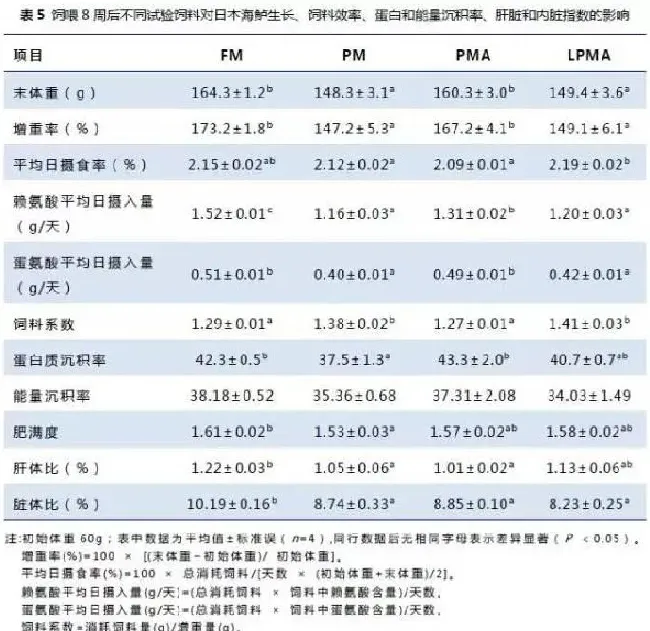
生长指标数据见表5。PMA组的增重率、饲料系数、蛋白沉积率和能量沉积率与对照无显著差异(P>0.05)。而PM和LPMA组的生长性能比FM和PMA组低(P<0.05)。LPMA组的平均日摄食率比PM组和PMA组高(P<0.05)。LPMA和PM组赖氨酸、蛋氨酸的平均日摄入量显著低于FM和PMA组,因此降低了LPMA和PM组的生长性能。鱼的增重率与赖氨酸和蛋氨酸的摄入量呈线性关系,分别为Y=33.169+260.81X,R2=0.571;Y=0.1154+940.13X,R2=0.714(图1)。与FM组相比,PM组的肥满度、脏体比和肝体比显著较低,PMA组的肝体比和脏体比显著较低,LPMA组的脏体比显著较低(P<0.05)。而LPMA和FM组的肝体比没有差异(P>0.05)。
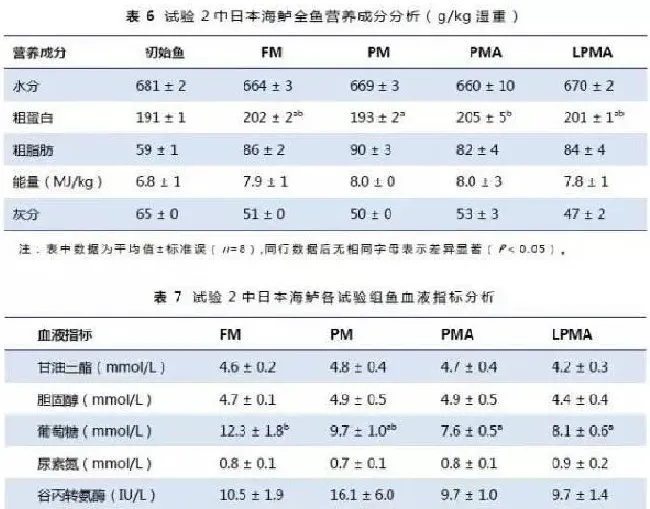
不同蛋白水平及鱼粉替代对全鱼水分、脂肪、能量和灰分没有影响,但PMA组全鱼蛋白含量显著高于PM组(表6)。不同饲料组血清中的谷丙转氨酶活性、甘油三酯、胆固醇和尿素氮含量没有差异(P>0.05,表7),而FM组血清中谷草转氨酶活性,谷草转氨酶/谷草转氨酶以及葡萄糖水平显著高于其它组(P<0.05)。
4、讨论
本研究表明饲料原料干物质表观消化率变化范围为68.7%(牛肉骨粉)~92.9%(猪肉粉),这主要是受饲料中灰分和纤维素含量的影响(Robaina等,1997;Bureau等,1999)。日本海鲈对各饲料原料中的蛋白质有较好的消化能力,这些结果与之前在其它肉食性鱼类中的研究结果相同(Gaylord等,1996;Laining等,2003;Zhou等,2004;Liu等,2009)。猪肉粉具有较高水平的蛋白(886g/kg)和较低水平的灰分(45g/kg),其蛋白表观消化率最高(95.9%)。其它鱼类上的研究表明,饲料蛋白水平和蛋白消化率呈正相关(Smith等,1995;McGoogan&Reigh1996;Liu等,2009)。本研究中,尽管牛肉骨粉的总磷含量最高,但其磷表观消化率却最低。而猪肉粉、水解羽毛粉和血粉的磷表观消化率极低,这些原料用作替代蛋白源时,必需补充有效磷。饲料中的磷以多种化学形式存在,包括骨磷、植酸磷、有机磷、磷酸二氢钙/钠/钾、磷酸氢钙等。HuaandBureau(2006,2010)建立了鲑类、鲤科和罗非鱼类对饲料原料的磷消化率的数学模型。鲑类和鲤科鱼类似乎不能利用植物性原料中的植酸磷,而罗非鱼能够利用部分植酸磷(大约27%)。本试验结果发现日本海鲈对动物性蛋白源(鱼粉、牛肉骨粉和鸡肉粉)及植物性蛋白源(豆粕)中的磷有较高的消化率(高于43.2%)。
尽管所有试验组都有较高的蛋白质消化率,但鱼粉、猪肉粉、喷雾干燥血粉和豆粕组总氨基酸表观消化率要高于牛肉骨粉、鸡肉粉、水解羽毛粉组。日本海鲈对不同蛋白源中氨基酸的消化率有所差异,因而在制作饲料配方时只是基于氨基酸的检测水平是不准确的。
必需氨基酸的缺乏被认为是限制鱼类对替代蛋白源利用的最重要的因素(Glencross等,2007)。与大多数价格低廉的蛋白源相似,猪肉粉中的限制性氨基酸为赖氨酸、蛋氨酸和苏氨酸,而且必需氨基酸和非必需氨基酸的比值(0.49)远远低于鱼粉中的数值(0.97)。
本试验结果显示猪肉粉饲料组中缺乏赖氨酸和蛋氨酸,结果导致此试验组鱼体增重率显著下降。PMA组的鱼体生长状况与鱼粉组相似,这表明日本海鲈与其它肉食性鱼类一样能够很好的利用晶体氨基酸。
许多研究采用理想蛋白的概念预测鱼类必需氨基酸的需要量(Wilson,2002;Mai等,2006;Peres等,2009)。从这些以及其它研究可以看出理想蛋白模型只是反映了对应鱼类品种的全鱼氨基酸组成(Brown,1995;Kaushik等,2008)。尽管如此,根据全鱼组成或者是肌肉组成计算出的必需氨基酸需要量还是被高估了(Mai等,2006;Kaushik等,2008)。与本试验结果类似的是在海鲷中的研究,试验结果显示在理想氨基酸模式下,饲料中只有三分之一的鱼粉能够被混合蛋白源(玉米蛋白粉、小麦面筋蛋白、去皮豌豆、菜粕)替代。作者得出结论,当饲料中鱼粉被替代后影响鱼体生长的主要因素是摄食量的显著减少(Gomez-Requeni等,2004)。Sheng(2009)在西伯利亚鲟鱼上的研究表明,在可消化理想蛋白模式下,混合动物蛋白源(牛肉骨粉,鸡肉粉,喷雾干燥血粉和水解羽毛粉)或混合植物蛋白源(豆粕、小麦面筋蛋白)都能够完全替代鱼粉,而且不影响生长。Kaushi等(2004)的研究发现用植物蛋白源完全替代欧洲海鲈饲料中的鱼粉时,最重要的是要保证每组饲料的营养素消化率以及各组鱼的摄食量相似。因此,为了更加精确地制作饲料配方并预测鱼体的生长状况,尤其是在考虑蛋白源替代和减少水产养殖废物排放时,需要同时考虑配方中的氨基酸推荐量和可消化氨基酸水平,而不仅仅关注饲料的总氨基酸含量,这一理念已经在畜禽动物和现代水产养殖中成功应用(Dari等,2005;LibaoMercado等,2006)。
降低饲料蛋白水平同时维持氨基酸的平衡,是维持畜禽正常生长的一种途径(Han等,1992;Hansen等,1993)。由于鱼类对碳水化合物利用能力低下,缺乏对氨基酸代谢的调控能力,因此鱼类与其它脊椎动物相比需要较高水平的蛋白用于维持生长和代谢(Cowey等,1989;Enes等,2009)。许多研究通过降低可消化蛋白水平,同时增加或不增加非蛋白能量水平来设计不同蛋能比梯度,以确定鱼类的最适氮能比(Cho等,1990;Cho等,2001)。Sheng(2009)研究发现在理想蛋白质模式下,饲料蛋白质水平从40%降到36%不会影响西伯利亚鲟鱼的生长,且显著减少氮排放。Yamamoto等(2005)的研究发现,在低蛋能比的饲料中添加需要量1.2倍的必需氨基酸后鱼体的生长没有受到影响,且提高了饲料氨基酸的利用率。但在本试验中,日本海鲈摄食根据理想蛋白模式设计、蛋白水平低于最适需要量的饲料后,生长显著受到影响,最重要的原因是必需氨基酸摄入量的显著减少。LPMA组中摄食量的增加并没有使日本海鲈的赖氨酸和蛋氨酸日摄入量达到需要量。与此同时,饲料中高水平的无氮浸出物降低了可消化能,进而也影响了日本海鲈的生长。
除了鱼体蛋白水平,各组饲料对全鱼其它组分基本上没有影响。日本海鲈摄食氨基酸平衡饲料,即使饲料蛋白降低,体成分也没有受到影响(LPMA组)。而猪肉粉组鱼体蛋白含量显著降低,饲料系数却显著较高,因此蛋白质沉积率也显著降低。大量研究表明,鱼类摄入缺乏必需氨基酸的高蛋白水平饲料将会增加环境中氮的排放(Kaushik等,2004;Sheng,2009;Zhao等,2010)。
尽管血液指标是检测蛋白质和脂类代谢的重要指标,但很少见到关于饲料中蛋白源影响日本海鲈血液指标的报道(Gomez-Requeni等,2004;Geurden等,2008)。本试验测定的血液指标(甘油三酯、胆固醇、葡萄糖、尿素氮、谷丙转氨酶和谷草转氨酶)结果与其它硬骨鱼类相似,如欧洲海鲈和鲤鱼(Lemaire等,1991;Svobodova等,2006),但谷丙转氨酶和谷草转氨酶活性显著低于阿穆尔河鲟鱼、中华鲟和大西洋鲑(Shi等,2006;Hemre等,2007)。此外,鱼体血液指标具有年度周期变化的特性且可能会受到细菌感染的影响(Acerete等,2009)。本试验结果发现,用猪肉粉替代鱼粉时显著降低鱼体血液中葡萄糖水平和谷草转氨酶活性。由于蛋白含量更高的蛋白源(猪肉粉)替代鱼粉时,多余的饲料配方空间将会被碳水化合物填充,碳水化合物水平的升高将会影响鱼体的蛋白质和糖类代谢。
本试验结果表明,在理想蛋白条件下,能够用115g/kg的猪肉粉替代饲料中160g/kg的鱼粉。日本海鲈摄食低蛋白水平(400.9g/kg)的饲料会抑制鱼体的生长,但其生长率与摄食蛋白水平较高(439.9g/kg)、氨基酸不平衡饲料的鱼体生长率近似。尽管各组饲料均按照理想蛋白模式配制,但是必需氨基酸缺乏(PM组)或者降低饲料蛋白水平(LPMA组),都不能满足日本海鲈达到最适生长的需求。(完)
(原文:WangJ,YunB,XueM,etal.Apparentdigestibilitycoefficientsofseveralproteinsources,andreplacementoffishmealbyporcinemealindietsofJapaneseseabass,Lateolabraxjaponicus,areaffectedbydietaryproteinlevels[J].AquacultureResearch,2012,43(1):117-127.)
1、来源:《中大水生通讯》第51期
2、作者:广州市诚一水产科技有限公司郭定乾/译
3、广州市诚一水产科技有限公司微信号:gzchengyi2013

























